March 2018 News
We finally get done with updating our web site, after several months of playing around with it, so we hope you like it!
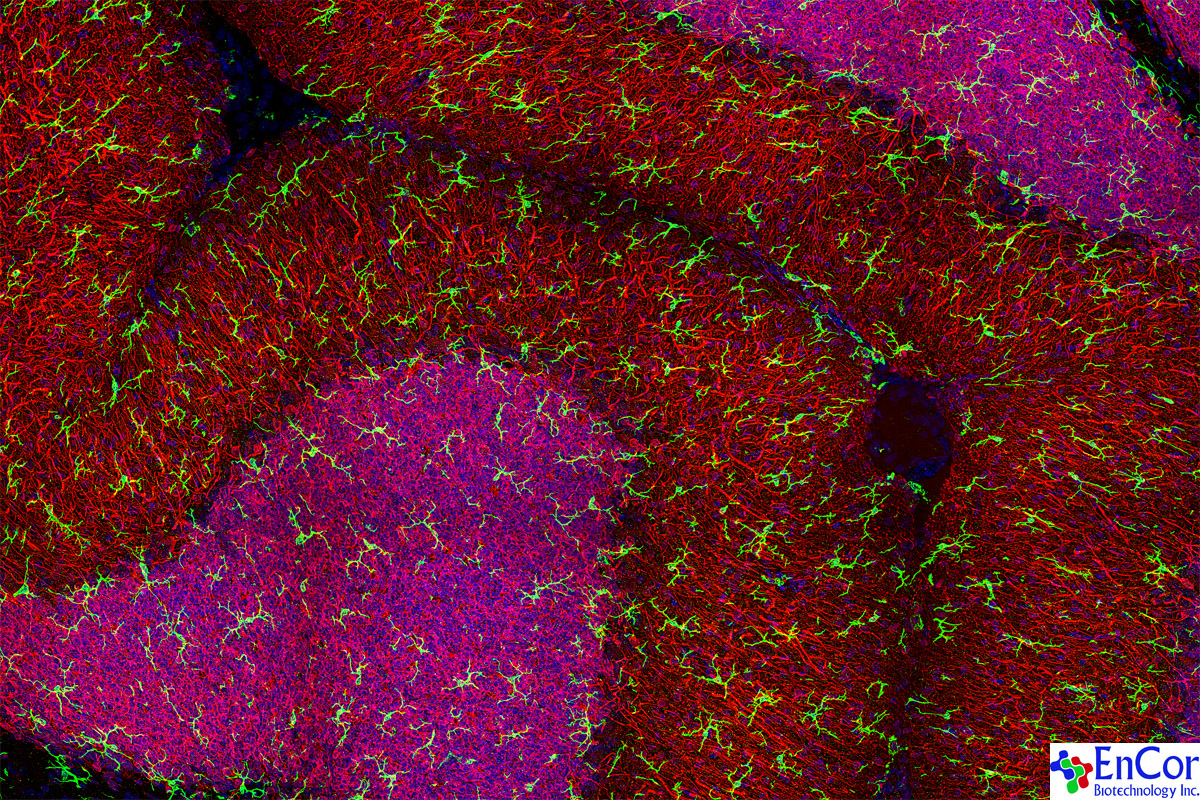
We have made an excellent rabbit polyclonal antibody to IBA1 (a.k.a. AIF1), RPCA-IBA1. The IBA1 protein is heavily expressed in microglia but not in neurons, astrocytes, oligodendrocytes or other kinds of CNS cell. As a result, antibodies to IBA1 are widely used to identify and study microglia in CNS cultures and sections. Microglia are the immune cells of the CNS and have important roles in the cellular responses to a variety of CNS damage and disease states. We also have an excellent high resolution poster showing staining of microglial cells with this antibody on rat cerebellum, counterstained withe the neuronal marker MAP2, see poster-27. Here is an image of that.
{kind=link}
We continue to characterize our antibodies in ever greater detail; for example we recently epitope mapped our two monoclonal antibodies to the intermediate filament protein vimentin, MCA-2A52 and MCA-2D1. This allows us to confidently predict which species these antibodies will work on. For another example see our new findings made on the widely used mouse monoclonal antibody to neurofilament NF-L MCA-DA2.